Breuer Lab










Papers & Presentations
Drag Forces and Unsteady Wakes Behind a Poro-Elastic Membrane Disk
Unsteady forces and flows behind a poro-elastic membrane disk
ResearchGate PaperDrag forces and unsteady wakes behind a poro-elastic membrane disk
ResearchGate PaperDiscover 2024: Drag Forces and Unsteady Wakes Behind a Poro-Elastic Membrane Disk
Coupled poro-elastic behavior of hyper-elastic membranes
Socha Lab






























Papers & Presentations
Tongue Behavior of Arboreal Colubrids During Gap Crossing
Tongue-sticking: Tongue behavior of arboreal colubrids during gap crossing
Zoe King, Sydney Haywood, Giovanni Morris, Jeffery Anderson-Jr, Joshua Pulliam, Jerry Wong, Beckett Socha, Ulmar Grafe, Salwa Khalid, Jake Socha
Tongue movement in snakes has been previously attributed to chemo- and mechanosensation characterized as a sweeping, vertical oscillation of the tongue, followed by the tongue being pulled back into the mouth to allow particle transfer to sensory organs. In previous studies, flying snakes (Chrysopelea) have been observed to protrude their tongue without oscillation in the context of locomotion. Do close relatives of flying snakes also exhibit this behavior? To investigate this question, we recorded the tongue behavior in close relatives (Ahaetulla prasina, Dendrelaphis pictus, caudolineatus, and formosis) as well as less related species (Dryocalamus subannulatus, Boiga cynodon, drapiezii, and nigriceps, Lycodon capucinus) as they traversed gaps. The setup consists of two artificial branches oriented horizontally and suspended 118 cm off the ground. Each snake completed four trials, recorded on four GoPro Hero4 cameras, crossing gaps of 0%, 15%, 30%, and 45% of the snake's snout to vent length. Among these species, only the close relatives of flying snakes (Ahaetulla and Dendrelaphis spp.) exhibited the presence of a tongue protrusion variant when crossing a gap. This behavior followed an oscillation, stick, oscillation (OSO) pattern. Generally, the tongue stick could be characterized by a rigid proximal half of the tongue while the tines show variable motion. The other species tested did not show any signs of tongue stick behavior. All snakes in this study were caught and examined at the University of Brunei Darussalam. This research was supported in part by the National Science Foundation (NSF) under grant numbers 1922516 and 2027523.
Body flattening in a close relative of flying snakes, Dendrelaphis pictus
Sydney Haywood, Zoe King, Giovanni Morris, Jeffery Anderson-Jr, Joshua Pulliam, Jerry Wong, Beckett Socha, Ulmar Grafe, Salwa Khalid, Jake Socha
Flying snakes (Chrysopelea) are the only snakes that have been observed to use dorsoventral flattening while locomoting: they expand their ribs, allowing for more surface area and enabling them to glide. However, not all species have been observed closely for the presence of this behavior. Recently, we observed possible dorsoventral flattening in a closely related species to flying snakes, specifically in Dendrelaphis pictus, commonly known as the painted bronzeback. Although it cannot glide, Dendrelaphis pictus is capable of a similar behavior to Chrysopelea: jumping. Does D. pictus use dorsoventral flattening when jumping? To determine if D. pictus utilize dorsoventral flattening, they were incentivized to jump using a horizontal gap-crossing setup, and recorded with multiple cameras to examine the body before they jumped. We found that the anterior body flattened in 4 specimens tested. To compare this behavior to rib movement during breathing, we also conducted recordings of the snake while breathing at rest in a tank. While D. pictus has been observed using dorsoventral flattening, it differs significantly to that of Chrysopelea: D. pictus only appear to pulsate in and out of dorsoventral flattening, and only in one part of the body. This could explain why Dendrelaphis are not able to generate sufficient aerodynamic forces to glide from tree to tree. This study demonstrates that some traits specific to gliding may have evolved prior to gliding in flying snakes. This research was supported in part by the National Science Foundation (NSF) under grant numbers 1922516 and 2027523.
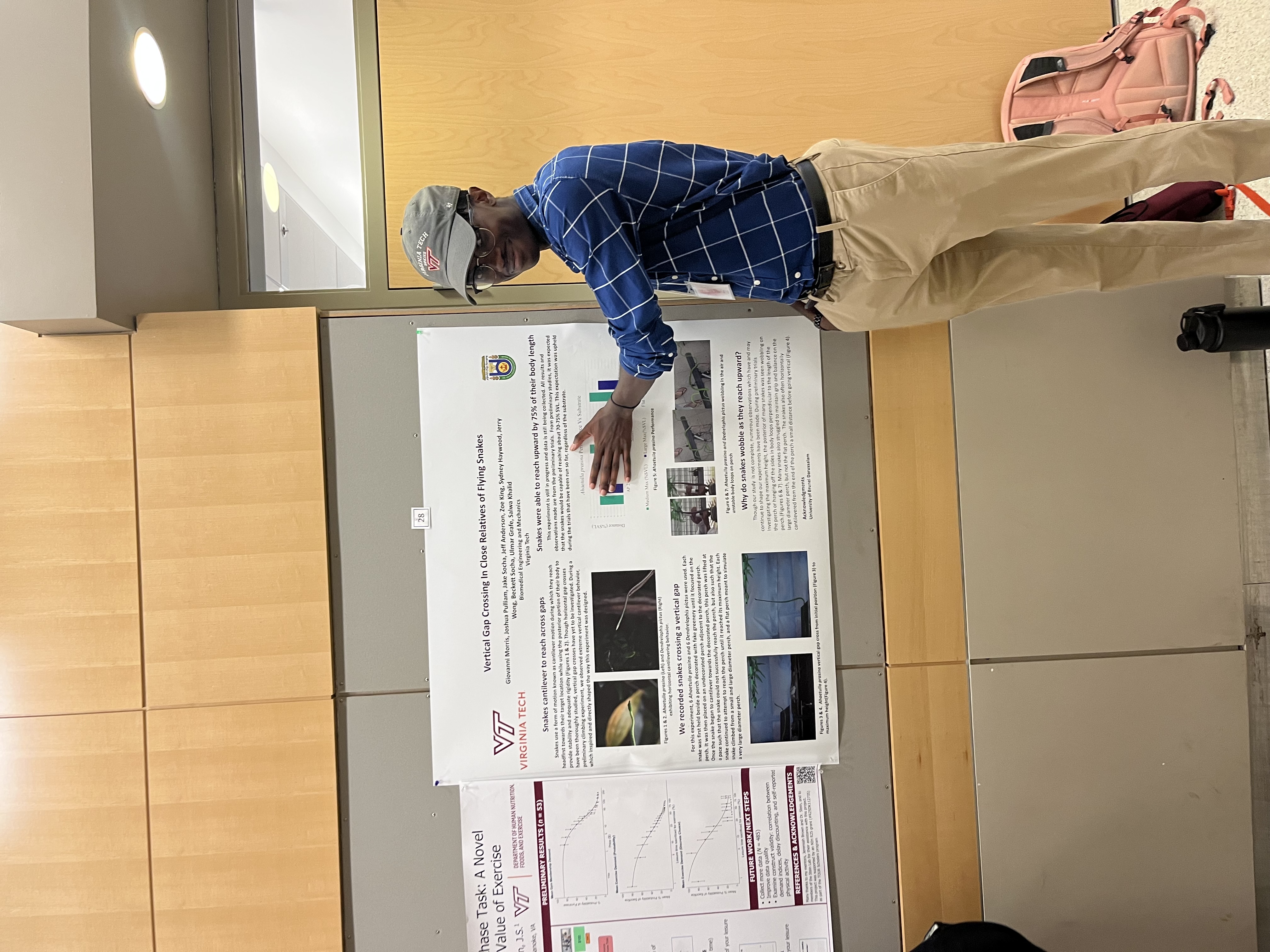
Vertical Gap Crossing in Close Relatives of Flying Snakes
Giovanni Morris, Sydney Haywood, Zoe King, Jeffery Anderson-Jr, Joshua Pulliam, Jerry Wong, Beckett Socha, Ulmar Grafe, Salwa Khalid, Jake Socha
Though limbed animals possess the ability to reach across horizontal and vertical gaps, limbless animals must find alternative methods of overcoming gaps in the arboreal environment. One strategy employed by snakes is cantilever locomotion. When navigating from one surface to another, snakes extend their bodies headfirst into the gap until they have reached their target. The snakes must provide proper stability and rigidity with the body in order to prevent themselves from buckling or pitching into the gap. How do snakes cope with gaps oriented in the vertical axis? Here, we investigated the vertical gap-crossing abilities of snakes, focusing on the effect of the substrate curvature on a snake's ability to cantilever. In this experiment, 6 painted bronzebacks (Dendrelaphis pictus) and 6 Asian vine snakes (Ahaetulla prasina) were allowed to cross from a base to a target perch, starting from one of four origin perches of varying curvature (diameter = 27, 43, and 157 mm, and a flat plane). The target perch was initially set at a small vertical distance (based on the size of the snake in percentage of snout-to-vent (SVL) length). As the snake rose to cross the gap, the distance was increased manually until the snake reached its maximum cantilever height and failed to reach the perch. Based on preliminary data, it is expected that the snakes will be able to gap cross at least 70-75% SVL. This research was supported in part by the National Science Foundation (NSF) under grant numbers 1922516 and 2027523.
SICB 2023: Tongue-sticking in arboreal colubrids during gap crossing
1505 Zoe King, Sydney Haywood, Giovanni Morris, Jeffery Anderson-Jr, Joshua Pulliam, Jerry Wong, Beckett Socha, Ulmar Grafe, Salwa Khalid, Jake Socha
Tongue-flicking in snakes is characterized by a rapid, high-frequency oscillation of the tongue, but other behaviors are known to be used. In tongue-sticking, the tongue is protruded statically without oscillation, a behavior previously noted in flying snakes (Chrysopelea) in the context of locomotion, specifically gap crossing and jumping prior to gliding. Is this behavior unique to flying snakes, or do other snakes also employ this form of tongue movement? And if so, how does the arboreal context influence the behavior? To investigate these questions, we recorded tongue behavior in sister taxa to Chrysopelea (Ahaetulla prasina, Dendrelaphis pictus, caudolineatus, and formosus) as well as outgroup species (Dryocalamus subannulatus, Boiga cynodon, drapiezii, and nigriceps, Lycodon capucinus) as snakes traversed a gap between two horizontally oriented perches. Snakes were first allowed to cross the perches in abutted position (a control, with no gap), and then were presented with gaps of 15%, 30%, and 45% SVL (snout-vent length) in random order. Among the species tested, only the close relatives of flying snakes (Ahaetulla and Dendrelaphis spp.) exhibited tongue sticking, present both in the control and gap trials. Furthermore, tongue sticking was more likely at the largest gap sizes, suggesting that tongue sticking plays a mechanosensory role associated with gap-crossing locomotion. This research was supported in part by the National Science Foundation under grant numbers 1922516 and 2027523.